Die Sprache der Pflanzen
Forschungsbericht (importiert) 2005 - Max-Planck-Institut für chemische Ökologie
Einleitung
Wie jeder lebende Organismus werden auch Pflanzen von ihrer Umwelt beeinflusst und sind in der Lage, auf äußere Reize zu reagieren. Neben zahlreichen abiotischen Faktoren wie z. B. Licht, Wasser, Strahlung oder Nährstoffe muss die Pflanze auch mit biotischen Einflüssen wie Konkurrenten, Pflanzenfressern (Herbivoren) und Pathogenen (Bakterien, Viren, Pilze) umgehen. Der Gefahr, die von Schädlingen ausgeht, sind Pflanzen trotz ihrer "Unbeweglichkeit" keineswegs hilflos ausgeliefert, denn sie besitzen zahlreiche Schutz- und Verteidigungsmechanismen, die entweder dauerhaft zum Einsatz kommen oder nur bei Bedarf genutzt (induziert) werden. Beispiele für kontinuierlich vorhandene Abwehrorgane sind Dornen, Stacheln oder Pflanzenhaare (Trichome). Sie dienen meist der physischen Abwehr von Schädlingen, indem sie die Pflanze für diese schwerer zugänglich machen. Darüber hinaus sind Pflanzen auch in der Lage, sich chemisch zu verteidigen, und zwar entweder direkt, zum Beispiel durch Giftstoffe, die für den Schädling toxisch sind oder dessen Verdauung stören, oder indirekt über die Abgabe von Duftstoffen, die z. B. Feinde des Schädlings anlocken. Die Produktion chemischer Abwehrstoffe ist für die Pflanze allerdings sehr aufwändig; um diesen Aufwand so gering wie möglich zu halten und somit Energie und Ressourcen zu sparen, wird die chemische Abwehr häufig nur bei Bedarf induziert.
Können Pflanzen sprechen und werden sie erhört?
Die bei Insektenbefall induzierte Abgabe verschiedener Duftstoffe kann der attackierten Pflanze also dazu dienen, räuberische Insekten zu Hilfe zu rufen, um die Population des Pflanzenschädlings zu reduzieren [1]. Das legt die Vermutung nahe, dass die Duftstoffe ebenso auf benachbarte, noch nicht attackierte Pflanzen wirken und diese vor der nahenden Gefahr warnen könnten, wobei sich die Frage stellt, ob und wie die Empfängerpflanze auf dieses „Warnsignal“ reagiert. Tatsächlich beginnt diese Reaktion bereits mit der Aufnahme des Duftstoffes an der Blattoberfläche, gefolgt von verschiedenen Schritten der Signalweiterleitung. Reaktionen der Empfängerpflanze auf der transkriptionellen Ebene der Gene lassen sich gut nachweisen, ebenso wie Änderungen im Proteom (Gesamtheit der Proteine) und im Metabolom (Gesamtheit der Stoffwechselprodukte). Daher beschränken sich auch viele der bisherigen Studien auf einen oder zwei dieser Bereiche. Dabei bleibt es jedoch meist fraglich, ob sich Duftstoff-induzierte Änderungen auf einer dieser Ebenen in der Fitness der Empfängerpflanze niederschlagen und ob die Wahrnehmung und Verarbeitung dieser durch Schädlinge induzierten pflanzlichen Signale eine ökologische Relevanz hat.
Experimentelle Schwierigkeiten
Erste Hinweise auf die Existenz einer Kommunikation zwischen attackierten und unbefallenen Pflanzen wurden schon vor ca. 20 Jahren gefunden. Baldwin und Schultz konnten 1983 zeigen, dass die Verwundung von Pappel- und Ahornpflänzchen die Produktion giftiger Phenole und Tannine induziert und dass in benachbarten, unverletzten Pflänzchen der Phenol- und Tanningehalt fast ebenso stark anstieg. Ähnliche Ergebnisse brachte eine Studie von Rhoades (1983) an Weiden (Salix sitchensis) hervor. In Fraßversuchen mit Raupen konnte gezeigt werden, dass die Blattqualität von attackierten Pflanzen geringer war als die von unverwundeten Pflanzen. Dieser Effekt war auch an unbefallenen Pflanzen nachweisbar, die neben angefressenen Bäumen standen, jedoch keinen Wurzelkontakt hatten. Die nahe liegendste Erklärung für diese Ergebnisse war eine Kommunikation zwischen verwundeten/attackierten Pflanzen und ihren unverletzten Nachbarn mithilfe flüchtiger organischer Substanzen (englisch: volatile organic compounds, kurz VOCs).
Die Ergebnisse dieser frühen Studien veranlassten viele Forschergruppen, sich dem Thema „Kommunikation zwischen Pflanzen über VOCs“ zu widmen. Die zahlreichen Forschungsarbeiten verdeutlichen allerdings auch die experimentellen Schwierigkeiten, die einen Nachweis des Phänomens einer pflanzlichen "Sprache" erschweren. In vielen Studien wurden die Pflanzen unrealistischen experimentellen Bedingungen ausgesetzt. Häufig wurden sie in luftdichte Behälter eingeschlossen [2], was mehrere Probleme mit sich bringt. Zum einen werden durch diesen Aufbau die Duftstoffe im Inneren der Kammer konzentriert, zum anderen wird der physiologische Status der Pflanze verändert: Sobald photosynthetische Prozesse ablaufen, sinkt die verfügbare Menge an CO2 in der Kammer, was zur Reduktion der Kohlenstoff-Fixierung führt. Um diesen Mangel zu kompensieren, können Pflanzen z. B. die Anzahl geöffneter Stomata (Spaltöffnungen) erhöhen. Dadurch werden jedoch die Mesophyllzellen verstärkt den Duftstoffen ausgesetzt und die möglichen Reaktionen einer Pflanze auf natürliche Duftstoffmengen amplifiziert oder verfälscht. Offene Systeme, die einen kontinuierlichen Luftfluss bieten, vermeiden diese Komplikationen.
Darüber hinaus wurden anstelle von intakten Pflanzen abgeschnittene Blätter oder Zweige als Duftproduzenten (Emitterpflanzen) oder als Duftempfänger benutzt. Mittlerweile ist aber bekannt, dass sich die Herbivoren-induzierte Duftstoffzusammensetzung abgeschnittener Blätter von der intakter Pflanzen unterscheidet. Durch Abschneiden entzieht man den Pflanzenteilen darüber hinaus wichtige Versorgungsgrundlagen, was ihren gesamten physiologischen Status verändert. Abgeschnittene Blätter oder Zweige sind daher für diese Untersuchungen nicht geeignet, weder als Duftproduzent noch als -empfänger.
Der Einsatz synthetischer Duftstoffe bietet die Möglichkeit, das von der Pflanze abgegebene VOC-Gemisch in seine Bestandteile zu zerlegen und die aktive(n) Substanz(en) zu identifizieren. Hier besteht die Schwierigkeit darin, die Pflanzen den natürlichen Mengen an Duftstoffen auszusetzen, Interaktionen zwischen mehreren Komponenten zu erkennen und die Schwankungen der Abgabe des Duftgemisches, die durch das Fressverhalten des Schädlings hervorgerufen werden, mit einzubeziehen. Freilandstudien ermöglichen realistischere experimentelle Bedingungen als Labor- oder Gewächshausversuche, wobei hier die Schwierigkeit darin besteht, die aktiven VOCs zu identifizieren.
Im Folgenden soll auf ein besonders intensiv untersuchtes Modellsystem eingegangen werden, welches gleichzeitig verdeutlicht, wie kompliziert und zeitintensiv konventionelle Freilandversuche sein können.
Wüstensalbei und wilder Tabak
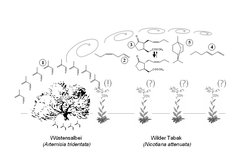
Karban et al. konnten im Jahr 2000 zeigen, dass Tabakpflanzen (Nicotiana attenuata), die neben verwundeten Wüstensalbei (Artemisia tridentata tridentata) gepflanzt wurden, deutlich weniger Herbivorenschaden erlitten und mehr Samenkapseln produzierten als Pflanzen neben unverwundetem Wüstensalbei. Nicotiana attenuata ist ein einjähriger Brandkeimer, der typisch für die Great Basin Wüste im Südwesten Nordamerikas ist. Diese Pflanze eignet sich besonders gut für Untersuchungen Herbivoren-induzierter Abwehr, da die zugrunde liegenden Mechanismen bereits ausgiebig untersucht wurden. Artemisia tridentata tridentata, ein mehrjähriger Busch, ist ein ebenso typisches Gewächs dieser Region, das gelegentlich auch in unmittelbarer Nachbarschaft des wilden Tabaks wächst (Abb. 1). Es produziert große Mengen des Phytohormons Jasmonsäure und dessen methylierten Derivats, welches in zwei verschiedenen Strukturformen (Epimeren) abgegeben wird (Abb. 2).
 und wilder Tabak (Nicotiana attenuata, im Vordergrund) sind typische Spezies der Great Basin Wüste im Südwesten Utahs, USA. Die Interaktion zwischen diesen beiden Arten ist derzeit das am besten untersuchte Modellsystem für den Informationsaustausch zwischen Pflanzen mittels oberirdischer Duftstoffe.")

 und Wüstensalbei (Artemisia tridentata tridentata). Pflanzt man Tabakpflanzen in die unmittelbare Nähe (15 cm Abstand) von verwundeten Wüstensalbeipflanzen, so werden sie weniger von Herbivoren geschädigt und produzieren mehr Samenkapseln als Tabakpflanzen, die neben unverwundetem Wüstensalbei wachsen. Verwundeter Wüstensalbei gibt eine Vielzahl stark flüchtiger (z. B. Methacrolein (1)) und weniger flüchtiger Duftstoffe (z.B. cis-3-Hexenal (2) und trans-2-Hexenal (4), oxygenierte Monoterpene wie Cineol (5), Thujon und Kampfer und zwei Epimere des Methyljasmonats (3)) ab. Während der Transport stark flüchtiger Duftstoffe eher über Diffusion erfolgt, werden die weniger flüchtigen Substanzen wahrscheinlich durch Luftturbulenzen fragmentiert und als Schwaden über größere Distanzen bewegt.")
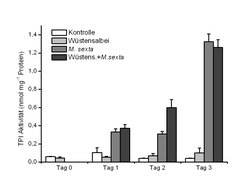
Nach Verwundung steigt die Abgabe des Methyljasmonats um das 9- bis 12fache [3, 4, 5]. Da Methyljasmonat als Pflanzenhormon schon seit langem bekannt ist, galt es für lange Zeit als der Kandidat für die aktive Substanz, die die Kommunikation zwischen Tabak und Wüstensalbei vermittelt. Experimente mit synthetischem Methyljasmonat konnten dies jedoch nicht bestätigen. Eine jüngere Studie der Jenaer Arbeitsgruppe zeigt, dass bestimmte Bestandteile des vom Wüstensalbei abgegebenen VOC-Gemischs (trans-2-Hexenal und Methacrolein) einen indirekten Effekt auf die benachbarten Tabakpflanzen haben. Die beiden Moleküle induzieren nämlich nicht direkt die Produktion von Abwehrsubstanzen (z. B. von Proteinase-Inhibitoren, die die Verdauung der Blattnahrung im Schädling hemmen), sondern verstärken bzw. beschleunigen deren Produktion nur im Falle eines Herbivorenbefalls ([6], Abb. 3).
 in wildem Tabak (Nicotiana attenuata) nach Empfang Verwundungs-induzierter Duftstoffe des Wüstensalbeis. Gezeigt ist TPI-Aktivität in unbeschädigten Tabakpflanzen (Kontrolle) und in Tabakpflanzen, die entweder dem Duftstoffgemisch des Wüstensalbeis für 24 Stunden ausgesetzt waren, die über 4 Tage von Raupen (Manduca sexta) angefressen wurden und die vor dem Raupenfraß den Duftstoffen des Wüstensalbeis ausgesetzt waren (Wüstens.+M.sexta). Die Werte von Tag 2 zeigen, dass vorherige Exposition zu den Duftstoffen des Wüstensalbeis die Raupen-induzierte TPI-Produktion beschleunigt.")
Diese durch den Wüstensalbeiduft vermittelte „Vorbereitung” der Tabak-Empfängerpflanze auf einen möglichen Insektenbefall steht also im Gegensatz zur direkten Induktion der Abwehr durch VOCs und scheint für die Empfängerpflanze die „sparsamere“ Variante zu sein. Um Abwehrmechanismen (z. B. die Produktion des giftigen Alkaloids Nikotin oder der Proteinase-Inhibitoren, die die Verdauung der Schädlinge stören) zu induzieren, muss die Pflanze Energie und Ressourcen investieren, die dann nicht mehr für andere wichtige Prozesse, wie Wachstum oder Fortpflanzung zur Verfügung stehen. Die direkte Induktion von Verteidigungsmechanismen durch Herbivoren-induzierte VOCs der befallenen Nachbarpflanze würde solche Kosten für die Empfängerpflanze und somit eine Reduktion ihrer Fitness verursachen, auch wenn sie am Ende gar nicht attackiert wird. Die Strategie der „Vorbereitung“ der Abwehrantwort verhindert dagegen unnötige Fitnesskosten, da ein erhöhter Verbrauch von Ressourcen und Energie nur im Falle eines tatsächlichen Schädlingsbefalls stattfindet. Dieses Phänomen konnte auch an Maispflanzen gezeigt werden, die nach Empfang von Duftstoffen in der Lage waren, ihre Herbivoren-induzierte Jasmonsäurebildung und Duftstoffabgabe zu verstärken (im Vergleich zu Herbivoren-befallenen Pflanzen, die die Duftstoffe nicht empfangen hatten [2]). Allerdings wurden in dieser Studie die Effekte der Duftstoffe auf benachbarte Pflanzen z. T. unter unrealistischen Bedingungen gemessen (geschlossene Kammer, abgeschnittenes Blattmaterial als Duftproduzent).
Ein neues eperimentelles Design ermöglicht realistische Laborbedingungen
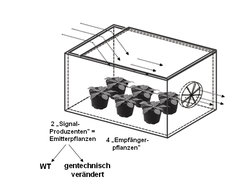
In einer kürzlich erschienenen Studie der Jenaer Arbeitsgruppe zu diesem Thema wird ein experimentelles Design vorgestellt, welches zwei Ansätze vereint und einige der oben genannten experimentellen Probleme vermeidet [7]. Zum einen wird ein offenes System verwendet, das die Emitter- und Empfängerpflanzen kontinuierlich mit Luft versorgt (Abb. 4).
. Frische Luft wird durch eine Öffnung auf der Oberseite der Box eingesogen. Als Duftstoffproduzenten (Emitterpflanzen) werden entweder Wildtyp-Pflanzen (WT) oder gentechnisch veränderte Pflanzen eingesetzt, denen bestimmte Bestandteile des Verwundungs- bzw. Herbivoren-induzierten Duftstoffgemisches fehlen.")
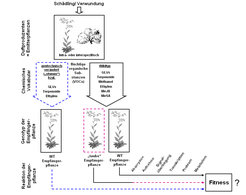
Darüber hinaus werden neben Wildtyp-Emitterpflanzen auch zwei gentechnisch veränderte Tabakpflanzenlinien eingesetzt, deren Herbivoren- bzw. Verwundungs-induziertem VOC-Gemisch bestimmte Bestandteile fehlen. Dies wird durch gezielte genetische Transformation erreicht, welche Proteine "stilllegt", die für die Biosynthese von Duftstoffen benötigt werden. Durch den Einsatz von transgenen Pflanzen, die in der Synthese von Duftstoffen bzw. Duftstoffgruppen verändert wurden – denen quasi ein Teil ihres chemischen „Vokabulars“ fehlt – ist es möglich, Effekte genau dieser Substanzen auf die Nachbarpflanzen zu untersuchen (Abb. 5). Die Vielzahl verschiedener verfügbarer Duftstoffmutanten und die Kenntnis der an der Biosynthese der VOCs beteiligten Gene ermöglichen eine umfassende Untersuchung des Schädlings-induzierten Duftgemisches und die Suche nach den aktiven Substanzen. Diese Studien können durch die Verwendung von Pflanzen, bei denen die Rezeption bestimmter Stoffgruppen ausgeschaltet ist (z. B. die Arabidopsis-etr1-Linie, die das volatile Pflanzenhormon Ethylen nicht erkennen kann und sozusagen „taub“ ist für Ethylen), vervollständigt werden.
 können Duftstoffen ausgesetzt sein, die entweder von schädlingsbefallenen Nachbarpflanzen der gleichen oder einer anderen Spezies (z. B Wüstensalbei) emittiert werden. Solche Duftstoffgemische bestehen unter anderem aus den Blattalkoholen und -aldehyden (englisch: green leaf volatiles, GLVs), Terpenoiden, Methyljasmonat, Methylsalicylat, Methanol und Ethylen. Verschiedene biotische Stressfaktoren beeinflussen dieses „chemische Vokabular“, das die Emitterpflanze produziert, in Bezug auf Qualität, Quantität und zeitliche Regulierung. Wird das Signal von der Empfängerpflanze detektiert, könnte sie auf der Ebene der Signaltransduktion, des Transkriptoms, des Proteoms und des Metaboloms darauf reagieren. Dies könnte letzten Endes in signifikanten Änderungen der Fitness resultieren. Der Vergleich der Antwort auf Wildtyp (WT)-Duftstoffe mit der Antwort auf teilweise „verstummte”, transgene Emitterpflanzen, die bestimmte Duftstoffe nicht mehr abgeben können, erlaubt die Identifizierung derjenigen Substanzen, die die Interaktion zwischen Emitter- und Empfängerpflanzen vermitteln. „Taube“ transgene Empfängerpflanzen, denen die funktionellen Rezeptoren für individuelle Duftstoffe fehlen, können die Studien zur Bioaktivität einzelner Duftstoffe vervollständigen.")
„Stumme“, transgene Pflanzen helfen, die „Duftstoffsprache“ zu übersetzen
Unter Verwendung des oben beschriebenen Versuchsdesigns überprüften die Wissenschaftler die biologische Aktivität der Blattalkohole und -aldehyde sowie terpenoider Duftstoffe (cis-α-Bergamoten) [7]. Dazu benutzten sie transgene Tabakpflanzen als Emitterpflanzen, denen entweder ein wichtiges Enzym (Hydroperoxidlyase) zur Synthese der Verwundungs-induzierten Blattalkohole und -aldehyde (as-hpl-Linie, die deutlich weniger Verwundungs-induzierte Blattalkohole und -aldehyde produziert) bzw. ein Enzym zur Synthese der Jasmonsäure (Lipoxygenase 3) fehlt, das nötig ist, um die Terpenemission zu induzieren (as-lox-Linie, die deutlich weniger Herbivoren-induziertes cis-α-Bergamoten produziert). In verschiedenen Experimenten wurde untersucht, ob Verwundungs- bzw. Herbivoren-induzierte VOCs von Wildtyp-Emitterpflanzen bestimmte Verteidungsmechanismen (die Produktion von Nikotin und von Proteinase-Inhibitoren (PIs)) oder die Synthese des dazugehörigen Signalmoleküls (Jasmonsäure) beeinflussen. Um die genannte „Vorbereitungs-Hypothese“ zu überprüfen, testete die Arbeitsgruppe ebenfalls, ob die Bildung von Nikotin, PIs oder Jasmonsäure nach Exposition und darauf folgender Verwundung der Empfängerpflanze in dieser verstärkt ist. Um einen eventuellen Effekt der VOCs auf eine spezifische Substanz(-gruppe) eingrenzen zu können, wurden in diesen Experimenten neben Wildtyp-Emitterpflanzen auch die zwei oben genannten, gentechnisch veränderten Tabak-Linien als Duftstoffproduzenten eingesetzt in der Erwartung, dass ihren Duftstoffgemischen die eventuell biologisch aktiven Substanzen fehlen und sie daher unfähig sind, die o. g. Effekte in den Empfängerpflanzen auszulösen. Diese Experimente zeigten jedoch, dass weder das vollständige Wildtyp-Duftgemisch noch die unvollständigen Gemische der gentechnisch veränderten Emitterpflanzen die getesteten Abwehrsubstanzen beeinflussen oder deren Produktion verstärken bzw. beschleunigen.
Um Hinweise auf andere mögliche Effekte zu erhalten, weiteten die Max-Planck-Forscher die Untersuchungen durch Verwendung von Mikroarrays auf die Ebene der Genexpression aus. Diese Tests bestätigten, dass Verwundungs- und Herbivoren-induzierte VOCs die Abwehrmechanismen der Empfängerpflanze nicht beeinflussen. Allerdings stellte sich auch heraus, dass deutlich mehr Gene in der Empfängerpflanze reguliert waren, wenn sie Duftstoffgemische ohne Blattalkohole und -aldehyde (emittiert von transgenen as-hpl-Pflanzen) empfangen hatte. Ein Teil dieser Gene war nicht mehr reguliert, wenn das unvollständige Duftstoffgemisch mit synthetischen Duftstoffen wieder vervollständigt wurde. Obwohl diese Gene keinem bestimmten Stoffwechselweg zugeordnet werden konnten, lassen diese Ergebnisse auf eine suppressive Wirkung der Blattalkohole und -aldehyde schließen.
Diese Ergebnisse verdeutlichen den Wert gentechnisch veränderter Pflanzen als Werkzeug zur Erforschung der Kommunikation von Pflanzen [8]. Darüber hinaus wird es nun möglich, die biologische Aktivität einzelner Duftstoffe bzw. Duftstoffgruppen in Freilandexperimenten, also unter den natürlichen wie allerdings auch komplexen Umweltbedingungen zu untersuchen.