Wie können Pflanzen blattfressende Insekten erkennen und abwehren?
Forschungsbericht (importiert) 2009 - Max-Planck-Institut für chemische Ökologie
Einleitung
Interaktionen zwischen Pflanzen und pflanzenfressenden Arthropoden haben die außergewöhnliche Vielfalt von Pflanzen ebenso wie deren vielfältiges Stoffwechselgeschehen evolviert. Pflanzen haben zudem ausgeklügelte molekulare Mechanismen entwickelt, mit denen sie Insekten (und andere Lebewesen) erkennen und auf die Ursache der Interaktion mit dem Insekt entsprechend reagieren können (vorteilhafte Wechselwirkung, z. B. Bestäubung; schädliche Wechselwirkung, z. B. Herbivorie). Da Insekten auch Überträger pathogener Mikroorganismen sind – die Erreger werden übertragen, während die Tiere an den Blättern fressen –, sind die Mechanismen, derer sich die Pflanzen bedienen, um Insektenfraß wahrzunehmen und darauf zu reagieren, wahrscheinlich komplex und überlappen möglicherweise mit den Mechanismen zu Wahrnehmung von und Reaktion auf Mikroorganismen.
Nicotiana attenuata, die Modellpflanze der chemischen Ökologie
Nicotiana attenuata ist eine einjährige Tabakpflanze, die in den Wüsten im Südwesten der USA heimisch ist. Sie wurde im Institut als Modellsystem etabliert, um die genetische Grundlage ökologischer Raffinesse mit besonderer Betonung der Pflanzen-Insekten-Interaktionen zu verstehen [siehe weiterführender Link Nr. 1]. Die Forschung zeigte, dass N. attenuata Blattfraß durch Insekten von einem rein mechanischen Schaden und einer Infektion durch Pathogene unterscheiden kann, was der Pflanze somit eine maßgeschneiderte Abwehrreaktion ermöglicht. Die Erkennung von Blattfraß durch Raupen erfolgt primär über die Perzeption von Komponenten aus den oralen Sekreten (OS) der Insektenlarven. Insbesondere Fettsäure-Aminosäure-Konjugate (FAC) in den oralen Sekreten von Raupen des Tabakschwärmers Manduca sexta sind notwendig und ausreichend, um in dieser Pflanze spezifische Reaktionen auf den Fraßschaden auszulösen [1]. Diese Tatsache sowie weitere Studien legen damit die Existenz von zentralen, durch Herbivorie aktivierbaren Regulatoren in den Blättern von N. attenuata nahe, die schon auf eine winzige Menge an FACs reagieren können. Wie FACs von der Pflanze erkannt werden, ist weitgehend genauso unbekannt wie die molekularen Mechanismen, die dem Auslösen spezifischer Abwehrreaktionen gegen Insekten zugrunde liegen.
Die durch OS und FACs ausgelösten Reaktionen in N. attenuata umfassen eine extensive Umprogrammierung ihres Transkriptoms, Proteoms und Metaboloms, um Abwehr- und Toleranzreaktionen hervorzubringen. Solche Umstellungen erfordern Energie und Ressourcen des Primärstoffwechsels und bewirken deshalb eine komplexe Reorganisation der Ressourcenverteilung in der Pflanze. Dazu gehören auch Veränderungen in der Photosyntheseleistung und in den Sink/Source-Verhältnissen innerhalb der Pflanze, also welche Blätter zum gegebenen Zeitpunkt Photoassimilate im Überschuss produzieren (Source) und welche Blätter Photoassimilate netto importieren (Sink), [2].
Eine beträchtliche Zahl der durch blattfressende Insekten induzierten Reaktionen beruht auf der Biosynthese von Molekülen der Jasmonatfamilie (JA). Diese von Lipiden abgeleiteten Signalsubstanzen spielen eine grundlegende Rolle bei der Auslösung pflanzlicher Resistenzreaktionen gegen beißende Angreifer. In N. attenuata wird die wundinduzierte JA-Produktion durch Zugabe von OS oder FACs verstärkt [1]. Die JA-Biogenese ist ein entscheidender Faktor für die pflanzliche Resistenz gegen Herbivoren, die Mechanismen jedoch, die der Aktivierung ihrer Biogenese unterliegen, sind immer noch weitestgehend unbekannt. Drei neuere Untersuchungen haben nun damit begonnen, die Wirkweise der durch FACs hervorgerufenen Stimulierung zu entflechten, um Informationen darüber zu liefern, wie 1) diese Elizitoren – so werden signalauslösende Moleküle genannt – die Aktivierung der JA-Biosynthese kontrollieren [3], 2) sie die Expression von früh regulierten Genen, die mögliche regulative Komponenten kodieren, beeinflussen und 3) wie FACs im verwundeten Blattgewebe verstoffwechselt werden, um neue aktive Signale zu generieren, die von der Pflanze wahrgenommen werden können.
Neue Einblicke in die Regulierung von Pflanzenreaktionen
I. Regulierung früher Mechanismen, die nach Auslösung durch FACs die JA-Biosynthese aktivieren
Eine kürzlich erschienene Studie [3] lieferte erste Hinweise auf die Beteiligung von zwei mitogenaktivierten Proteinkinasen (SIPK und WIPK) und dem NPR1-Regulator bei der Aktivierung von frühen Enzymreaktionen des JA-Biosyntheseweges nach FAC-Auslösung in N. attenuata-Blättern. Die detaillierte Analyse der frühen Veränderungen (innerhalb weniger Minuten) von Gehalten der Jasmonat-Ausgangssubstanzen, zusammen mit der Identifizierung einer plastidären Glycerolipase (GLA1), die für die De-novo-Biosynthese von JA in dieser Pflanzenart wesentlich ist, legt nahe, dass nach Verwundung und FAC-Auslösung ein rascher und spezifischer Fluss von freier α-Linolensäure in den JA-Biosyntheseweg stattfindet. Die Ergebnisse deuten weiterhin an, dass α-Linolensäure via Lipoxygenase-3 (LOX3) verstoffwechselt wird und somit wahrscheinlich eine enge physische Verbindung zwischen den Enzymreaktionen von GLA1 und LOX3 in N. attenuata existiert. Sie entspricht vermutlich denjenigen Mechanismen, die in Säugetieren bei der Aktivierung der Leukotrien-Biosynthese (Prostaglandine) beteiligt sind (Abb. 1), [4, 5].
. Die Erkennung dieser Moleküle durch die Pflanzenzellen löst Signale aus, die differentiell durch die Aktivierung von Glycerolipase A1 (GLA1) in Nicotiana attenuata eine Freigabe von 18:3-Fettsäuren aus Membranlipiden bewerkstelligen. SIPK und NPR1 nehmen an der Aktivierung dieses enzymatischen Schrittes teil, allerdings sind auch unbekannte und unabhängige Mechanismen beteiligt. Im Gegensatz dazu zielen WIPK-vermittelte Mechanismen auf die Aktivierung der OPDA-Biosynthese durch die Regulierung von Allenoxid-Synthase (AOS) und/oder Allenoxid-Zyklase (AOC). Wegen der chemischen Instabilität des AOS-Endprodukts EOT (12,13-Epoxy-18:3; OPDA: (9S,13S)-12-Oxo-Phytodiensäure) wird angenommen, dass die Enzyme AOS und AOC in engem Kontakt stehen.")
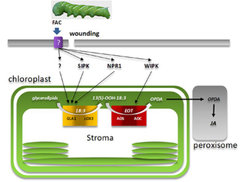
Der FAC-vermittelte vermehrte Einstrom von α-Linolensäure für die JA-Biosynthese via GLA1-Aktivierung hängt von SIPK und NPR1 ab, während WIPK bei der Kontrolle derjenigen Biosynthesestufen wirksam ist, die 13S-Hydroperoxy-Linolensäure in 12-Oxo-Phytodiensäure (OPDA) umwandeln, und zwar via Allenoxid-Synthase (AOS) und/oder Allenoxid-Zyklase (AOC; Abb. 1). Von WIPK, SIPK und NPR1 nimmt man an, dass sie extraplastidäre Proteine sind und deshalb zusätzliche Komponenten benötigt werden, um die aktivierenden Signale in das Stroma der Chloroplasten zu befördern, wo sich die Enzyme befinden, die die Initialstufen der JA-Biosynthese katalysieren (Abb. 1). Die weitere Identifizierung von Signaltransduktionskomponenten und die Untersuchung des Auslösens der Mechanismen, die die Enzymaktivitäten beeinflussen, wird entscheidende Informationen zum Verständnis darüber liefern, wie primäre Stresssignale in die Aktivierung von JA-Biosynthese-Enzyme übersetzt werden.
II. Regulatoren früher Abwehr- und Toleranzreaktionen in Nicotiana attenuata
SuperSAGE ist ein Verfahren zur absoluten Quantifizierung des mRNA-Gehalts, das in Kombination mit Next Generation Sequencing (NGS) eine unverfälschte und gründliche Analyse von Veränderungen im Transkriptom eines gegebenen Organismus ermöglicht. Die Wissenschaftler am MPI für chemische Ökologie nutzten dieses Verfahren, um die frühen transkriptionellen Veränderungen zu quantifizieren, die durch das FAC N-Linolenoyl-Glutaminsäure (18:3-Glu) in N. attenuata ausgelöst werden. Ziel ist es, mRNAs zu identifizieren, die regulierende Komponenten kodieren, also rare Transkripte mit sehr rascher FAC-ausgelöster Kinetik. Von 12744 identifizierten spezifischen Transkriptsequenzen wurden 547 innerhalb von 30 Minuten nach 18:3-Glu-Applikation unterschiedlich reguliert. Wichtig dabei war, dass die meisten Veränderungen bei Transkriptklassen auftraten, die mit mRNAs in geringen Konzentrationen korrelierten, wie z. B. eben jene, die üblicherweise regulierende Komponenten kodieren. Folgerichtig entsprachen mehr als 25% der 547 unterschiedlich regulierten mRNAs den mutmaßlichen regulierenden Komponenten, darunter unter anderem 30 Transkriptions-Regulatoren und 22 Proteinkinasen (Abb. 2).
. A So genannter „Volcano-Plot“, der den Log2 (fold-change: x-fache Veränderung; F (FAC): elizitiert, vs. W (Wounding): verwundetes Blattmaterial) versus Log10 (P-value: probability: Signifikanzwert) von 547 exprimierten mRNAs (identifiziert in Form von so genannten UniTags) zeigt. B Fold-change (F vs. W) – Verteilung der 547 unterschiedlich exprimierten mRNAs versus Häufigkeit. Die Punkte unterhalb der gestrichelten Linie entsprechen mRNAs mit geringer Häufigkeit (< 1000 Kopien pro Million). C Mittlere Masse (+/- Standardfehler) von M. sexta-Larven nach 4, 7, 11 und 15 Fütterungstagen mit Pflanzen, deren LPP-kodierendes Gen (Lipid-Phosphat-Phosphatase) ausgeschaltet worden war; EV (empty vector)-Pflanzen dienten als Kontrolle (n=32 für jeden Genotyp). D Mittlere Masse (+/- Standardfehler) von M. sexta-Larven nach 4, 7, 11 und 15 Fütterungstagen an EV-Pflanzen (Kontrolle) und Pflanzen, deren UnkA-kodierendes Gen (kodiert ein Protein mit unbekannter Funktion) ausgeschaltet worden war (n=32 für jeden Genotyp).")
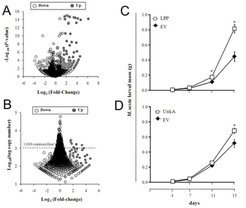
Einige dieser regulatorischen Kandidatengene wurden durch Virus Induced Gene Silencing (VIGS) funktionell charakterisiert, einem Verfahren, das auf das Ausschalten bestimmter Gene abzielt und somit eine Analyse der Pflanzenreaktion in Abwesenheit der Expression dieser Gene erlaubt. Die Analyse konnte zwei potenzielle regulierende Gene identifizieren – eine putative Lipid-Phosphat-Phosphatase (LPP) und ein Protein mit unbekannter Funktion (UnkA)– , die beide eine entscheidende Rolle bei der Regulierung der Interaktion zwischen M. sexta und N. attenuata spielen. Bei M. sexta-Raupen, die an Pflanzen gefressen haben, bei denen die Expression dieser Gene ausgeschaltet worden war, war die Gewichtzunahme signifikant höher im Vergleich zu Raupen, die an Kontrollpflanzen fraßen (Abb. 2). Somit hat diese Studie eine Grundlage für die Identifizierung von Regulatoren der pflanzlichen Abwehr- und Toleranzreaktionen geschaffen.
III. Schneller Stoffwechsel von FACs in verwundeten Nicotiana-attenuata-Blättern
. 13-LOX: 13-Lipoxygenase.")
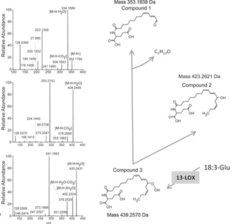
Um die Mechanismen besser zu verstehen, die der FAC-Erkennung durch Pflanzen zugrunde liegen, wurde die Verstoffwechselung eines wichtigen FACs aus den Oralsekreten von M. sexta, N-Linolenoyl-Glutaminsäure (18:3-Glu), direkt bei Kontakt mit verwundeten Blattoberflächen von N. attenuata-Blättern analysiert. 70% des synthetischen 18:3-Glu, auf verwundete Blätter aufgetragen, wurde innerhalb von 30 Sekunden metabolisiert. Dieser hitzelabile Prozess resultierte nicht in einer 18:3-Fettsäure und der Aminosäure Glutamat, also einer einfachen Spaltung des FAC, sondern in Metaboliten, die mehr oder weniger polar waren, verglichen mit 18:3-Glu: Tatsächlich waren 13-Hydroxy-18:3-Glu, 13-Hydroperoxy-18:3-Glu und 13-Oxo-13:2-Glu die am häufigsten gebildeten Metabolite, die mithilfe von LC-MS/MS (Flüssigchromatographie/Massenspektrometrie) identifiziert werden konnten (Abb. 3). Die Bildung dieser Metabolite war abhängig von der Aktivität der Lipoxygenasen 2 und 3. Pflanzen, bei denen die Expression der zugehörigen Gene ausgeschaltet worden war, zeigten niedrigere Raten einer 18:3-Glu-Oxidation und akkumulierten nur kleine Mengen von 18:3-Glu-oxidierten Derivaten im Vergleich zu Kontrollpflanzen. Ähnlich wie 18:3-Glu löste auch 13-Oxo-13:2-Glu eine verstärkte Produktion von JA in N. attenuata-Blättern aus, während dies bei 13-Hydroxy-18:3-Glu nicht der Fall war. Dies weist darauf hin, dass die Modifizierung von 18:3-Glu sowohl einen aktiven Elizitor als auch ein im Zusammenhang mit der JA-Biosynthese inaktives Molekül erzeugt. Möglicherweise geht die Erkennung von Elizitoren, die vom Insekt aus dessen Oralsekreten stammen, mittels oder nach Modifizierung durch Pflanzenenzyme vonstatten. Dies würde bedeuten, dass eine durch Pflanzen katalysierte Umwandlung von Insektenelizitoren, die in der Biogenese neuer, abgewandelter Elizitoren resultiert, eine weitere Ebene der Komplexität bei den Interaktionen zwischen Pflanzen und Insekten hinzufügt und neue Fragen aufwirft.
Perspektiven
Das Forschungsfeld der Pflanzen-Insekten-Interaktionen ist ein sich rasch entwickelndes multidisziplinäres Gebiet. Besonders die Verbindung der molekularen und ökologischen Expertisen ermöglicht einen leistungsstarken Ansatz zum besseren Verständnis von Genfunktion und Evolution vor einem ökologischen Kontext. Darüber hinaus ist das molekulare Verständnis von Pflanzen-Insekten-Interaktionen auch von grundlegender Bedeutung für die Generierung neuer Ansätze für die Bekämpfung von Schädlingen an Nutzpflanzen. Die molekulare Analyse ökologisch relevanter Interaktionen zwischen N. attenuata und seinen natürlichen Feinden auf verschiedenen Ebenen – von der Zelle bis zum Organismus – trägt bereits vielversprechende Früchte, die zu einem besseren Verständnis der Interaktionen zwischen Pflanzen und pflanzenfressenden Arthropoden beitragen.