Wie Insektenfraß Blätter zum Leuchten bringt
Forschungsbericht (importiert) 2015 - Max-Planck-Institut für chemische Ökologie
Calciumionen (Ca2+) sind wichtige interzelluläre sekundäre Botenstoffe in Signalnetzwerken von Pflanzen. Nach Fraßschäden öffnen sich spezifische Ionenkanäle und es kommt zu einem schnellen, vorübergehenden Anstieg des cytoplasmatischen Ca2+-Levels. Diese erhöhten Levels können mit biolumineszentem Aequorin verfolgt werden: Es bindet spezifisch Ca2+ und beginnt zu leuchten. Dieses Signal wandert mit ca. 1 bis 2 cm/min durch das direkt verbundene vaskuläre System und korrespondiert mit der Ausbreitungsgeschwindigkeit von Raupenfraß-induzierten elektrischen Signalen.
Pflanzen und ihre Verteidigung gegen Herbivore
Als nicht-mobile Organismen sind Pflanzen wie kaum eine andere Organismengruppe Änderungen in ihrer unbelebten (abiotischen) und belebten (biotischen) Umwelt unmittelbar ausgesetzt. Um ihr Überleben zu sichern, müssen sie auf solche Änderungen entsprechend reagieren. So bedingt Lichtmangel phototropes, auf das Licht gerichtetes Wachstum, Wassermangel den Verschluss der Schließzellen, hohe UV-Belastung die Synthese von Schutzpigmenten wie Flavonoiden, um nur einige Beispiele zu nennen. Als Primärproduzenten, die am Anfang jeder Nahrungskette stehen, sind Pflanzen neben abiotischen Stressfaktoren auch vielen biotischen Stressfaktoren, sprich Lebewesen ausgesetzt. Hier sind vor allem phytopathogene Mikroorganismen wie Pilze, Oomyceten und Bakterien aber auch Pflanzenviren zu nennen. Darüber hinaus existiert eine Vielzahl von Fraßfeinden, von Vertebraten über Arthropoden bis hin zu Nematoden. Insbesondere innerhalb der Arthropoden (Gliederfüßer) sind hier die pflanzenfressenden Insekten zu nennen, die die zahlenmäßig größte Gruppe der herbivoren Organismen repräsentieren.
Pflanzen und Insekten koexistieren nunmehr seit dem Paläozoikum; das heißt seit ca. 480 Millionen Jahren müssen sich Pflanzen gegen herbivore Insekten erwehren (Abb. 1). Das tun sie unter anderem durch die Synthese und Akkumulierung von toxischen, abschreckenden oder unverdaulichen Metaboliten, die alle der Verteidigung dienen. Mehr als 200.000 verschiedene chemische Substanzen, die zu verschiedenen Stoffklassen gehören und deren Wirkungen auf unterschiedlichen Mechanismen beruhen, sind mittlerweile bekannt [1]. Viele dieser Metaboliten sind konstitutiv vorhanden; andere sind induzierbar und werden nur bei Bedarf gebildet. Letzteres erfordert das effiziente Erkennen einer Herbivoren-Attacke (Reizerkennung), im Falle von Insektenfraß eine Kombination aus Verwundung und chemischen Signalstoffen aus dem Insektenspeichel, und eine schnelle Weiterleitung (Signaltransduktion) die letztlich zur Aktivierung und Etablierung adäquater Abwehrreaktionen gegen den Aggressor führt (zelluläre Reaktion). Dies kann zum Beispiel durch eine veränderte Regulation des Stoffwechsels und/oder der Genexpression erfolgen [1, 2].
.")

Calcium – ein intrazelluläres Signal
Neben anderen Signaltransduktions-Komponenten ist das Calciumion (Ca2+) schon seit langem als wichtiger intrazellulärer sekundärer Botenstoff in Zellen bekannt und somit auch für Pflanzenzellen von essenzieller Bedeutung, um auf Veränderungen in ihrer Umwelt reagieren zu können [3]. Dies betrifft das abiotische ebenso wie das biotische Umfeld und schließt Angriffe durch Pathogene oder Herbivoren mit ein [2]. In pflanzlichen Zellen ist die freie cytosolische Ca2+-Konzentration mit 100 nM (nanomolar) im Vergleich zum Apoplast (Zellwandbereich) oder der Vakuole (≥ 1 mM) sehr niedrig. Ausgelöst durch vielfältige äußere Reize kann in der Zelle ein schneller, vorübergehender Anstieg der cytoplasmatischen Ca2+-Konzentration um mehrere Größenordnungen als intrazelluläres Signal erfolgen. Dies geschieht durch das Öffnen von spezifischen Ionenkanälen. Um die Ca2+-Homöostase wieder herzustellen, wird nach wenigen Minuten der Basislevel der Ca2+-Konzentration im Cytoplasma wieder eingestellt. Parameter wie die Dauer, Intensität und intrazelluläre Verteilung der Calciumsignale, sowie das Muster von eventuellen Wiederholungen erlauben es, den äußeren Signalen/Reizen entsprechende Ca2+-Signale zuzuordnen [4]. Dies reicht allerdings bei weitem nicht aus, um die enorme Spezifität der nachgeschalteten Reaktionen auf die auslösenden Signale zu erklären. Weitere Elemente sind deshalb notwendig, die das Calciumsignal dekodieren können. Die erhöhten Konzentrationen an Calciumionen werden in der Zelle von spezifischen Ca2+-bindenden Proteinen wahrgenommen oder sie regulieren Kanäle, die den Endpunkt des Signalweges darstellen. Hierbei können Calciumionen auf zwei unterschiedliche Weisen als Signalmolekül fungieren: direkt und indirekt. Entweder besitzen Proteine – wie viele Proteinkinasen – spezifische Bindungsstellen für Ca2+ und die Aktivität dieser Kinasen wird durch die Ca2+-Bindung direkt beeinflusst, oder spezifische Proteine binden Ca2+, verändern daraufhin ihre Konformation und wechselwirken dann direkt mit Zielproteinen, um deren Aktivitäten zu regulieren. Calmoduline oder die pflanzenspezifischen calmodulin-like-proteins (CMLs) sind Beispiele für solche Calcium bindenden Proteine [5].
Calcium – ein interzelluläres, systemisches Signal?
Viele Erkenntnisse über die Rolle von Calciumionen in der Reizweiterleitung wurden mithilfe von gentechnisch veränderten Organismen erzielt. Hierfür machte man sich die Eigenschaften eines biolumineszenten Proteins zunutze, Aequorin, das mit hoher Affinität Ca2+ binden kann. Aequorin ist ein Photoprotein aus der Qualle Aequorea victoria, das im aktiven Zustand ein Luciferin als Cofaktor kovalent gebunden hat. Nach Bindung von Ca2+ emittiert es Lichtquanten. Die Intensität der Lichtemission korreliert mit der Calciumkonzentration und lässt somit deren Berechnung in jeweils dem Zellkompartiment zu, in dem das Aequorin exprimiert wurde, in der Regel im Cytoplasma [6].
Nahezu unbekannt war bislang die Rolle von Calciumionen in systemischen Reaktionen in von Insekten befallenen Pflanzen. Nachdem gezeigt worden war, dass in Arabidopsis thaliana ein durch Verwundung induziertes elektrisches Signal von Blatt zu Blatt wandert [7], lag die Vermutung nahe, dass auch Calciumionen eine Rolle in der systemischen Antwort auf Herbivorie spielen, obwohl Ca2+ selber eher immobil ist [3]. In Kooperation mit Kollegen der Universität Halle-Wittenberg haben Mitarbeiter der Abteilung Bioorganische Chemie des MPIs für chemische Ökologie diese Hypothese überprüft. Unter Verwendung von transgenen, Aequorin-exprimierenden A. thaliana-Pflanzen und Larven des afrikanischen Baumwollwurms, Spodoptera littoralis (Abb. 1), untersuchten sie deshalb in einem nicht-invasiven Ansatz unter Verwendung einer hoch-sensitiven Kamera die Ca2+-induzierte Lichtemission [8].
Der Raupenfraß an einem vorher definierten Blatt ließ als unmittelbare Reaktion der Pflanze angefressene Blattbereiche aufleuchten (Abb. 2, 0-5 min). Darüber hinaus konnte mit diesem Ansatz erstmalig sichtbar gemacht werden, dass das Calciumsignal tatsächlich systemisch auftritt und innerhalb von wenigen Minuten von einem befallenen Blatt in benachbarte Blätter wandert (Abb. 2). Dort kann das Calciumsignal entsprechende Abwehrreaktionen auslösen. Die kalkulierte Geschwindigkeit für dieses Signal beträgt 1 bis 2 cm/min, was mit der Geschwindigkeit übereinstimmt, die für die bei Raupenfraß-induzierten elektrischen Signale bestimmt wurde [7].

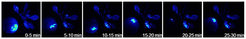
Interessanterweise war das Calciumsignal nicht in allen systemischen Blättern nachweisbar. Nur in den Blättern, die direkt über das vaskuläre System der Pflanze miteinander verknüpft waren, konnte das Leuchten nachgewiesen werden. Dieses Ergebnis kann dahingehend interpretiert werden, dass sich das mobile, durch Insektenfraß ausgelöste Signal im vaskulären System ausbreitet und nur in den direkt verknüpften Blättern ein Calciumsignal auslösen kann [8]. Was das mobile Signal tatsächlich ist, ein elektrisches oder doch ein chemisches Signal, muss noch aufgeklärt werden.
Fazit
Die Studie veranschaulicht, dass das nach Insektenfraß für das Auslösen pflanzlicher Abwehrreaktionen essenzielle Calciumsignal in seiner räumlichen und zeitlichen Ausbreitung in der gesamten Pflanze visuell verfolgt werden kann. Darüber hinaus konnte sie demonstrieren, dass es möglich ist, die Rolle des Botenstoffs Ca2+ in Pflanzen in verschiedenen physiologischen und ökologischen Zusammenhängen direkt in intakten Pflanzen zu untersuchen und so besser zu verstehen.
Literaturhinweise
Trends in Plant Science 20, 83-489 (2015)